- Home
- News
- Spotlight on Science
- Muscle as a shock...
Muscle as a shock absorber
31-03-2014
Muscle is the organ that generates force and movement in vertebrates and invertebrates. It also works as a sophisticated shock absorber that helps to absorb the energy of an impact, such as when landing from a jump or running. These so-called “eccentric contractions”, during which muscle fibres lengthen against the contractive force, are known to require less energy than for a concentric muscle action in which the muscle shortens as it contracts. The molecular mechanism underlying this function was investigated by means of high-resolution, low-angle diffraction experiments on muscle fibres. The results show that the braking mechanism of muscle is an intrinsic property of the muscle machinery consisting of the actomyosin system of interdigitating protein filaments found in all muscles.
Thick myosin filaments and thin actin filaments form precisely repeating arrays, called sarcomeres, along the muscle length, with tens of thousands of sarcomeres arranged in series in each skeletal muscle cell. In mammals, sarcomeres extend over a range of about 2 µm. The helical packing of actin and myosin filaments within the sarcomere gives rise to a rich low-angle diffraction pattern. During active contraction, the globular heads of myosin molecules, which protrude from the thick filaments at regular intervals, cyclically bind to, and release from, actin filaments, generating a sliding movement of the thick filaments relative to the thin filaments and shortening of the sarcomeres. This process requires hydrolysis of ATP, the fuel for the molecular motors. The actomyosin nano-machinery itself cannot lengthen the sarcomeres. Lengthening of the muscle and sarcomeres is achieved by contraction of the opposing muscle on the opposite side of the limb or by an external force.
Research using a double ergometer, a contraption in which two bicycles were connected back to back so that one rider was able to resist the pedalling of another rider, found that a slight rider was easily able to slow down and stop a much more powerful rider pedalling forwards [1]. The braking effort of the slight rider is an eccentric exercise and requires about four times less energy than that required to pedal forward against the same resistance, as measured by the amount of oxygen consumed. Eccentric exercise is beneficial to muscle growth and repair, and to alpine skiers [2] whose sport involves much eccentric contractions, for example in giant slalom turns [3].
During forced extension of active muscle (stretch), the utilisation of ATP decreases virtually instantly upon application of the stretch [4]. Stiffness of muscle also increases virtually instantly [5] indicating that the number of myosin motors bound to actin increases by a factor of nearly two, compared to the number of myosin molecules attached when the muscle is exerting maximum force with no shortening (isometric contraction).
In experiments performed at beamline ID02, the low-angle X-ray diffraction patterns obtained during isometric contraction of mammalian muscle fibres at in vivo temperature were directly compared with the corresponding patterns during stretch. Figure 1 depicts the ‘difference diagram’ obtained by the subtraction of the normalised isometric and stretch patterns and it reveals the extent to which stretch transforms the structural arrangement within the sarcomeres. Surprisingly, an increase in the number of myosin heads bound to actin – evident from the increase in muscle stiffness – was not accompanied by an increase in the intensities of the actin layer lines, A1 and A2, as one would have expected. Indeed, firm binding of myosin heads to actin would increase the electron density of an actin-based periodicity and should intensify the actin-based reflections. Conversely, the observed decrease during stretching shows that fewer myosin heads have the actin-based symmetry than during isometric contraction. These data are interpreted in terms of changes in the orientation and mode of attachment of myosin molecules to actin [6]. During contraction, myosin binding to actin proceeds from a non-stereo-specific binding mode to a stereo-specific mode, prior to detaching and re-attaching further along the actin filament.
 |
|
Figure 1. The normalised low-angle diffraction pattern during stretch (left), isometric contraction (centre) and their difference diagram (right). The difference diffraction diagrams collected from 9 runs of the protocol in 5 bundles of muscle fibres in the first experimental series. White and black correspond to an increase and decrease in the intensity during stretch compared to isometric contraction, respectively. X-ray reflections of interest are marked. |
During stretch however, the process is reversed. The stereo-specifically bound heads are pulled back towards their non-stereo-specific attachment mode as illustrated in Figure 2. Myosin molecules are constrained to 'walk' backwards along the thin filaments, maximising the actin-attachment time, which maintains the high stiffness that protects bone and joints from damage, and minimising, or even halting the utilisation of ATP. The stretching force unlocks the myosin heads that are stereo-specifically bound to actin, preventing them from consuming ATP. Stretching also promotes non-stereo-specific attachment of detached myosin heads, which stiffens eccentrically contracting muscle and absorbs mechanical energy, thus acting as an effective shock absorber.
 |
|
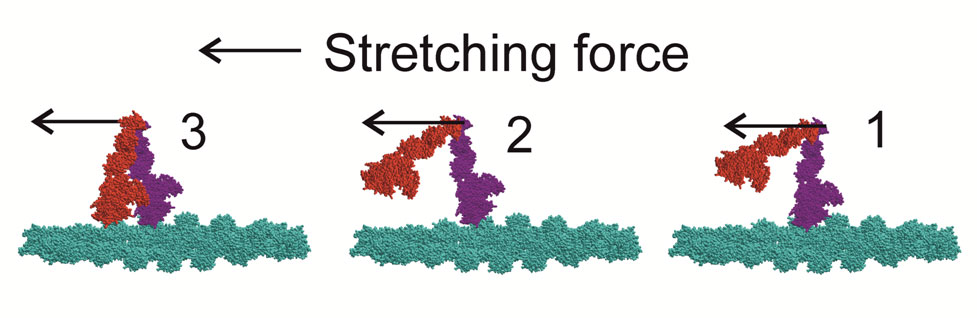
Figure 2. Schematic model of the movement of myosin heads upon muscle stretching that explains the data. During isometric contraction (state 1) only one (purple) head of a majority of myosin molecules is stereo-specifically bound to actin (cyan). Stretching unlocks the bound head to a non-stereo-specifically attached state and brings the distal part of the partner head to a position from which it can easily bind to a neighbouring actin monomer (state 2). Then the second (red) head quickly binds actin also non-stereo-specifically (state 3). As both heads are bound non-stereo-specifically, the A1 intensity is low and stiffness is high. Further stretching leads to detachment of the purple head followed by its rapid rebinding to an actin monomer 5.5 nm closer to the M-line of a sarcomere, thus producing ‘head over head’ walking. The M-line of the sarcomere is on the left and the Z-disk is on the right.. |
Principal publication and authors
M.A. Ferenczi (a,b), S.Y. Bershitsky (c), N.A. Koubassova (d), G.V., Kopylova (b), M. Fernandez (e), T. Narayanan (e), A.K. Tsaturyan (d), Why muscle is an efficient shock absorber, PLoS ONE, 23 January 2014.
(a) Molecular Medicine Section, NHLI, Imperial College London (UK)
(b) LKC Medicine, NTU (Singapore)
(c) Institute of Immunology and Physiology, Ural Branch RAS, Yekaterinburg (Russia)
(d) Institute of Mechanics, Lomonosov Moscow University (Russia)
(e) ESRF
References
[1] B.C. Abbott, B. Bigland and J.M. Ritchie, The physiological cost of negative work, J. Physiol. 117, 380 (1952).
[2] M. Gross, F. Lüthy, J. Kroell, E. Müller, H. Hoppeler, and M. Vogt, Int. J. Sports Med. 31, 572 (2010).
[3] H.E. Berg, O. Eiken, and P.A. Tesch, Med. Sci. Sports Exerc. 27, 1666 (1995).
[4] D.C. Bickham, T.G. West, M.R. Webb, R.C. Woledge, N.A. Curtin, and M.A. Ferenczi, Biophys. J. 101, 2445 (2011).
[5] E. Brunello, M. Reconditi, R. Elangovan, M. Linari, Yin-Biao Sun, T. Narayanan, P. Panine, G. Piazzesi, M. Irving, and V. Lombardi, PNAS 104, 20114 (2007).
[6] M.A. Ferenczi, S.Y. Bershitsky, N. Koubassova, V. Siththanandan, W.I. Helsby, P. Panine, M. Roessle, T. Narayanan, and A.K. Tsaturyan, Structure 13, 131 (2005).
Top image: Strong binding between myosin (red and purple) and actin (cyan) occurs when muscle acts as a shock absorber.
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.