- Home
- News
- Spotlight on Science
- Molecular form of...
Molecular form of copper in root cells of a non-hyperaccumulating plant grown in an urban setting
18-08-2014
Metal acquisition or detoxification in the roots of vascular plants occurs through biochemical mechanisms that involve specialised metal-complexing molecules. Current knowledge is largely based on hyperaccumulating species having high metal concentrations and may not apply to the most common plants in the environment. The molecular form of copper in the root cell walls of a non-accumulating species grown naturally in an urban setting was determined using a combination of X-ray fluorescence and absorption techniques at the nano-, micro-, and macroscopic scales.
Copper is an important metal for plant growth because it can be a nutrient or a toxin depending on its cellular concentration. Although all plants require copper, knowledge of its biochemistry in roots is based on relatively rare metal-accumulating species in highly contaminated natural or laboratory environments [1-2]. The roots of common native plants grown in an urban stormwater runoff basin provide a model for characterising the molecular mechanisms of metal uptake in non-accumulators at metal concentrations of concern to regulating agencies.
Metals enter plant roots along with water and move through the outer cortical region to the central stellar cells (Figure 1a). The biomolecules that transport, store, or sequester copper are considered to involve oxygen, nitrogen, or sulphur ligands [1]. It has been shown that copper is accumulated within cell walls rather than in cell vacuoles, but the precise identity of the ligands and details of their molecular structures have only been inferred [3,4]. We examined naturally grown field pennycress (Thlaspi arvense) from a stormwater runoff basin east of Lyon. This edible plant is sensitive to copper and has already been used in comparative studies with a model zinc and cadmium hyperaccumulating plant (Thlaspi caerulescens).
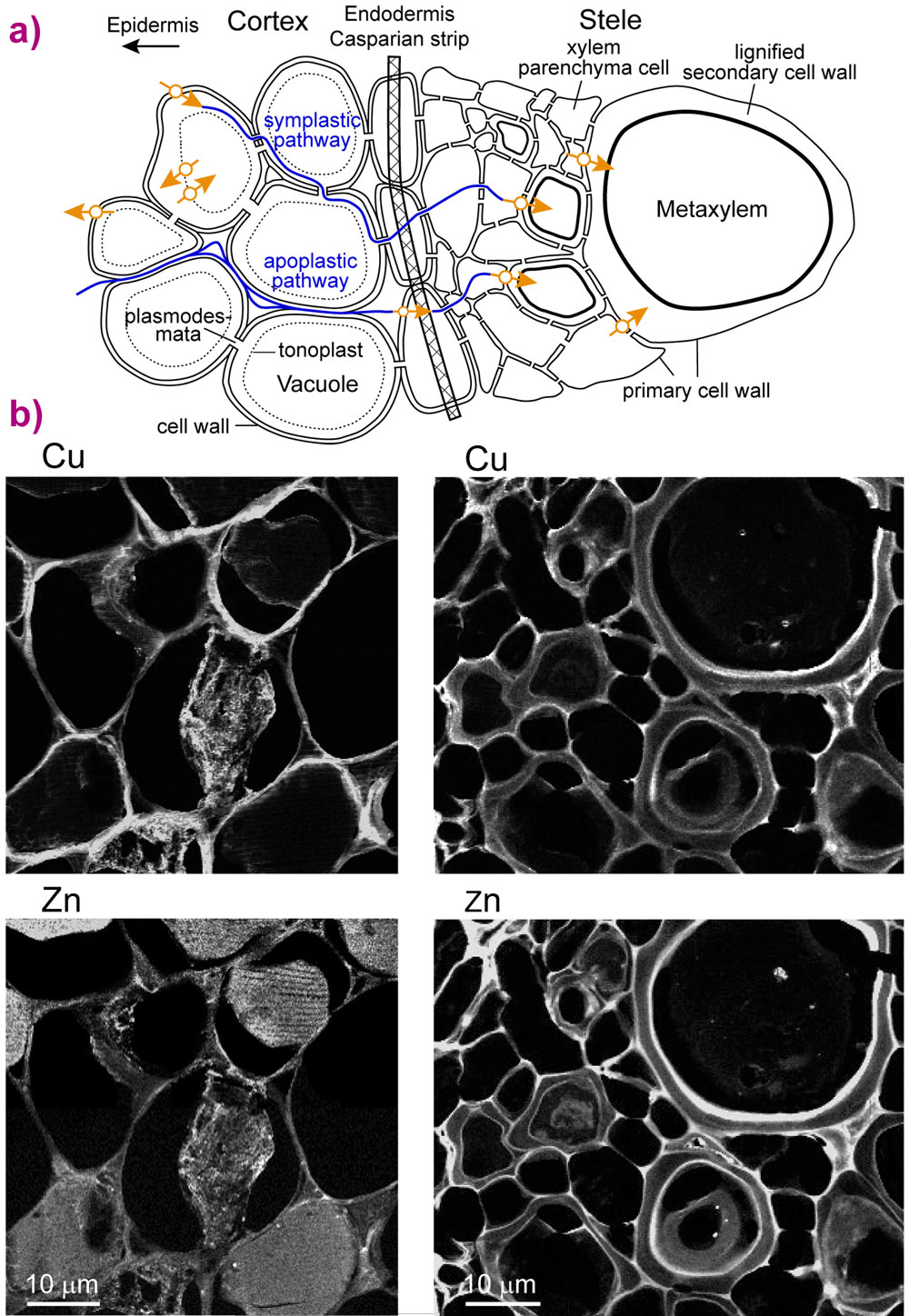 |
|
Figure 1. a) Sketch of a transverse root section showing the movement of metals around protoplasts (apoplastic path) and from cell to cell via plasmodesmata (symplastic path). The cell wall accumulates metals from the environment both during diffusive apoplastic flow of water and active removal from the protoplast to avoid toxicity. Orange arrows indicate specific protein transporters involved in Cu uptake and homeostasis in cytoplasm. b) Grayscale nano X-ray fluorescence images of Cu and Zn in the cortex (left) and stele (right) of a secondary root showing the compartmentation of Cu in cell walls and of Zn in cortical vacuoles and cell walls. Less than 5% of the copper is in vacuoles. The pixel size is 180 (V) x 180 (H) nm2 for a beam size of 69 (V) x 77 (H) nm2, FWHM. |
We found that copper in secondary roots (68 ± 25 mg Cu/kg dry root) is seven times more concentrated than in woody primary roots and exceeds cellular requirements. Nano-X-ray fluorescence images of cells in secondary roots frozen under in vivo conditions were obtained at ESRF beamline ID22NI (now ID16NI). The images confirm that copper is situated in cell walls in both the cortex and stele, unlike zinc, which was also found in vacuoles (Figure 1b). Micro-X-ray absorption spectra, measured at beamline ID22 and Advanced Light Source beamline 10.3.2, match spectra for copper complexed with the amino acid histidine in a bis- chelate structure (Figure 2). This cell wall complex was confirmed as being the dominant form of copper from bulk spectra of rapidly frozen fresh roots, obtained at ESRF beamline BM30B.
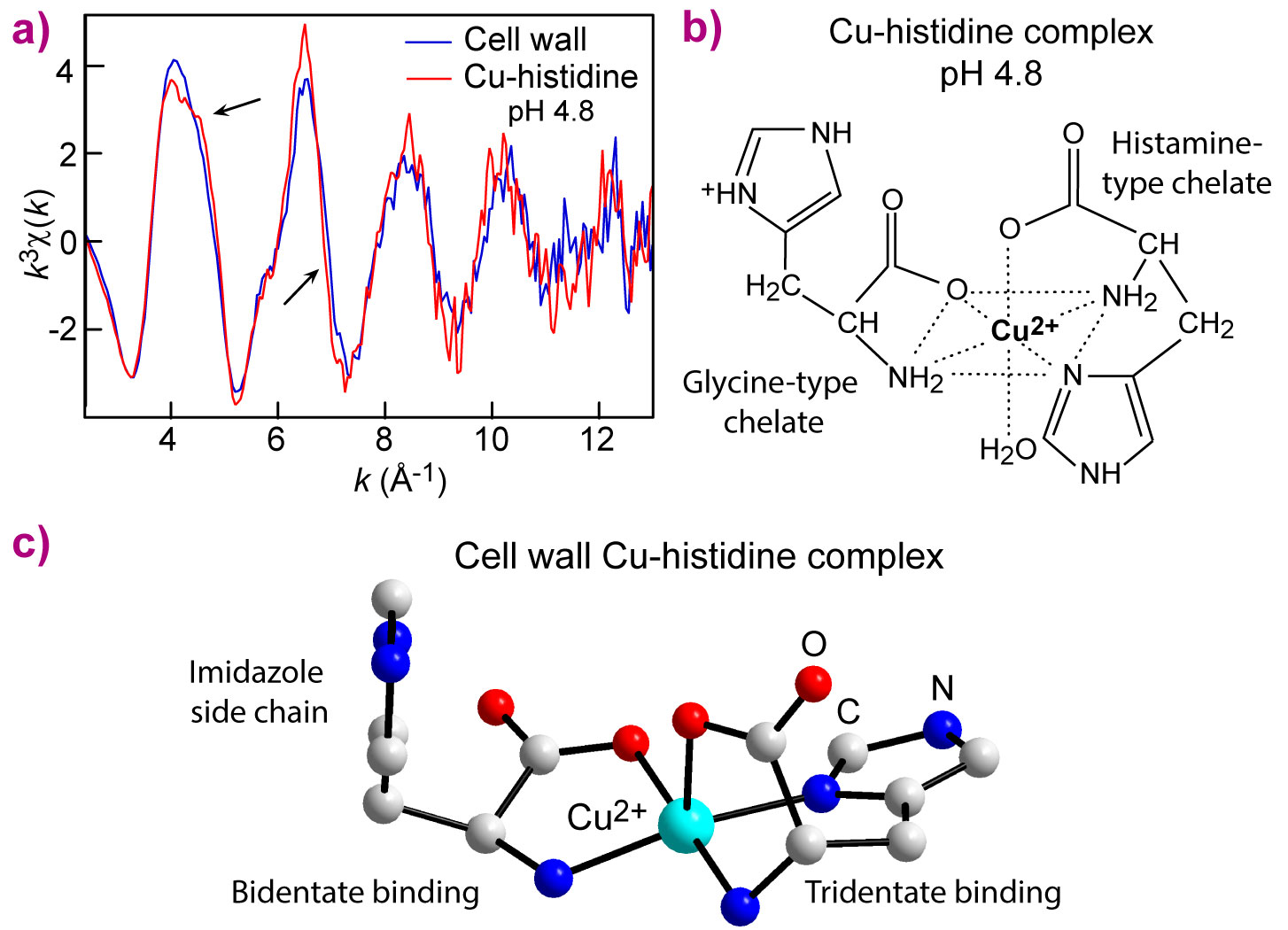 |
|
Figure 2. a) Copper speciation by Cu K-edge EXAFS spectroscopy. The cell wall spectrum closely resembles the spectrum of a Cu-histidine complex at pH 4.8, and distinctly differs from those of other Cu complexes with amino acids (not shown). b) The Cu-histidine complex has a bis-chelate structure formed from a five-membered glycine-like (gly) bidentate chelate and a six-membered histamine-like (him) tridentate chelate. c) The model for Cu complexation on cell walls is a five-coordinate square-pyramid, which differs from the Cu-histidine complex by the loss of the opposite water molecule. |
We propose that the copper-histidine complex has unique properties that could either stabilise copper in the cell wall or facilitate its mobility through the cell wall and into the cells depending on the amount of copper in the soil and the nutrient requirements of the plant. One L-histidine residue forms three bonds (tridentate) with copper and the other forms two bonds (bidentate) (Figure 2c). Tridentate binding would be stronger and could be part of a detoxification mechanism that reduces the cellular concentration of copper cations by stabilising the metal in the cell walls. Bidentate binding with a pendant imidazole side chain could allow hydrophobic interactions with proteins that transport copper through the cell walls into the root cells and from there into the growing plant.
This first report of metal imaging in cell walls at high resolution provides new information on copper acquisition in plants grown in soils at metal concentrations characteristic of urban environments. With advances in X-ray imaging and spectroscopy, identification and characterisation of the molecular structure of metal species at low concentration in other cellular regions of vascular plants such as the stem will become possible.
Principal publication and authors
Thlaspi arvense binds Cu(II) as a bis(L-histidinato) complex on root cell walls in an urban ecosystem, A. Manceau (a), A. Simionovici (a), M. Lanson (a), J. Perrin (a), R. Tucoulou (b), S. Bohic (b,c), S.C. Fakra (d), M.A. Marcus (d), J.-P. Bedell (e), K.L. Nagy (f), Metallomics 5, 1674-1684 (2013).
(a) ISTerre, CNRS and University of Grenoble Alpes, Grenoble (France)
(b) ESRF
(c) Grenoble Institut des Neurosciences, Grenoble (France)
(d) Advanced Light Source, Berkeley (USA)
(e) CNRS and University of Lyon, Lyon (France)
(f) University of Illinois at Chicago, Chicago (USA)
References
[1] H. Küpper et al., Plant Physiol. 151, 702-714 (2009).
[2] N. Verbruggen et al., New Phytol. 181, 759-776 (2009).
[3] J. Shi et al., Plant Soil 302, 163-174 (2008).
[4] J. Song et al., Biol. Plant 57, 581-586 (2013).
Top image: Nano X-ray fluorescence images of copper in the cortex (left) and stele (right) of a secondary root in field pennycress (Thlaspi arvense).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.