- Home
- News
- Spotlight on Science
- RNA virus replication...
RNA virus replication into ribonucleoproteins and its regulation elucidated from bunyavirus polymerase structure
24-08-2015
The structural characterisation of the La Crosse bunyavirus polymerase by X-ray crystallography and cryo-electron microscopy provides a detailed description of how bunyavirus polymerase can specifically recognise the viral RNAs, how the viral RNA genome can allosterically regulate the polymerase activity and the RNA traffic inside the polymerase during RNA synthesis. These discoveries explain many biochemical observations accumulated over decades of bunyavirus research, providing a detailed mechanistic understanding of bunyavirus RNA synthesis. Some of these findings can be extended to all negative stranded RNA viruses, such as influenza, Lassa and Ebola viruses.
In 1943, a new virus was isolated from Aedes mosquitoes during an investigation of yellow fever in the Semliki forest, Uganda. This virus, called Bunyamwera (BUNV), was the first of a new family of segmented negative stranded viruses that took the name Bunyaviridae. Since then, more than 350 strains of bunyavirus have been isolated around the globe and classified into five genera: Orthobunyavirus, Hantavirus, Nairovirus, Phlebovirus and Tospovirus, including many arthropod and rodent born human pathogens. La Crosse bunyavirus (LACV) belongs to Orthobunyavirus and is a potentially serious, but rare, mosquito-transmitted human pathogen that causes 50–100 cases of encephalitis per year in the USA, mainly affecting children [1].
Bunyavirus, in common with Arenavirus (e.g. Lassa virus) and Orthomixovirus (e.g. influenza virus), have their genomic RNA divided into several fragments that are coated by viral nucleoproteins and assembled with their polymerase to form circular RNA structures. This assembly, called the ribonucleoprotein (RNP), is the functional unit of replication and transcription [2]. Transcription is carried out by RNPs through a unique mechanism known as “cap snatching”, where 5’-CAP labelled cellular mRNAs are cleaved by a specific enzymatic domain located in the polymerase [3], and subsequently used for priming the viral mRNA synthesis. These functional similarities contrast with the sequence divergence between these viral polymerases. For example, the influenza virus polymerase is a heterotrimer [4] instead of a large single polymerase, also called the L protein, utilised by Arenavirus and Bunyavirus.
The structure of the LACV Orthobunyavirus polymerase [5] was obtained by protein crystallography using data collected at beamlines ID23-1 and ID29 at the ESRF, and cryo-electron microscopy using the EM platforms at IBS (PSB) and EMBL Heidelberg (Figure 1). The structure reveals several domains extending from the universally conserved polymerase core that form a large and complex protein fold where four access channels converge on a central polymerase reaction chamber (Figure 1). The bunyavirus polymerase reveals a striking structural similarity when compared with the recently reported structure of the influenza heterotrimeric polymerase [4], also determined using data collected at the ESRF. Therefore, this overall fold structure is conserved among all segmented negative stranded viruses, despite the complete lack of sequence homology. A recently reported electron microscopy structure of the vesicular stomatitis virus (VSV) also shows that the non-segmented negative strand viral (nsNSV) polymerases have a similar globular core to bunyavirus and influenza polymerases [5].
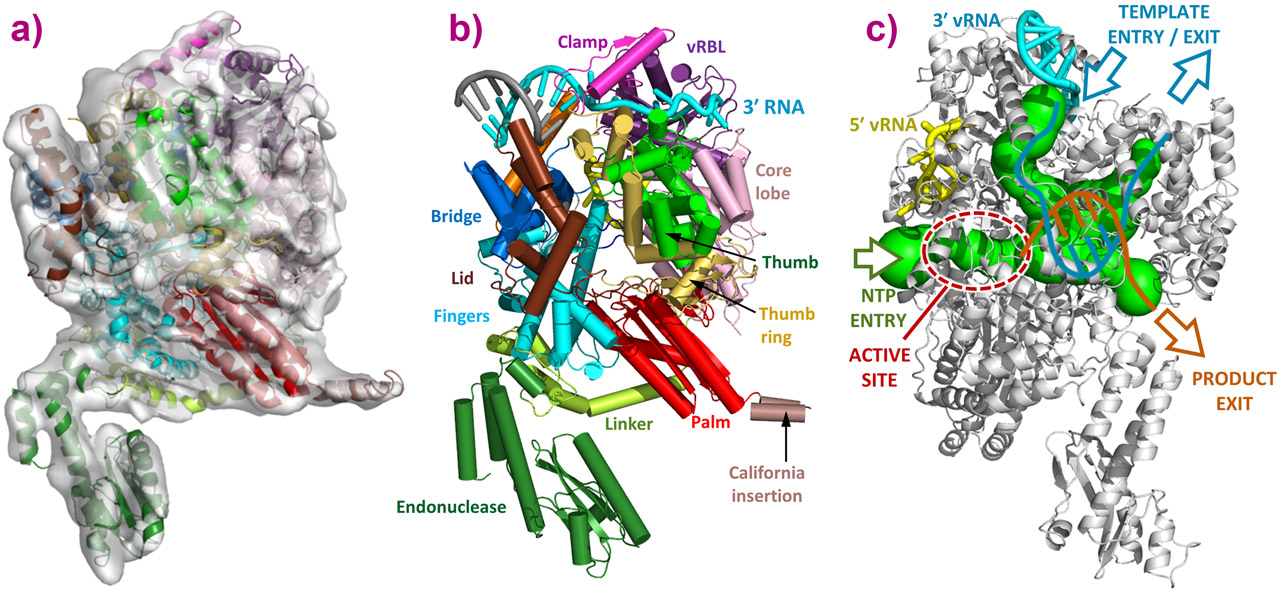 |
|
Figure 1. a) 3D reconstruction of the LACV polymerase apo-L1750 containing the entire dataset of cryo-EM imaged particles, determined at 8.3 Å resolution. b) Representation of the crystal structure of L1750 in complex with the 3’ (cyan) and 5’ (yellow) vRNA. b) A model of the elongation state showing trajectories of template RNA (cyan) and product RNA (orange) and NTPs through the polymerase tunnels (green). After formation of a short template-product duplex, accommodated in the interior cavity, template and product RNA strands exit separately along different tunnels, the template back towards the polymerase entry, where it can re-integrate into the RNP and the nascent strand to the bottom, where product processing occurs. |
The crystal structure of the L protein in complex with the viral RNAs clearly shows how the 3’ and 5’ RNA ends are bound at separate binding sites, and illustrates how the circularity of the RNP assembly is achieved (Figures 2 and 3). The high specificity for the self-viral RNA recognition is mediated by extensive sequence specific protein-RNA contacts into positively charged clefts outside the catalytic site, and close to the template entrance channel. The binding of the 5’-vRNA end has an allosteric effect on the active site that activates the polymerase by ordering the important ‘fingertips’ structural element in an active conformation (Figures 2 and 3).
 |
|
Figure 2. Representation of the LACV polymerase (grey) showing the disposition of key structural elements (arch, clamp, a-ribbon, fingertips, fingernode). The 5’ and 3’ vRNA extremities are coloured in yellow and cyan, nucleotide 11 in each case is coloured in red to highlight their wide separation (>20 Å). The figure shows the different binding sites for each RNA, and the impossibility of formation of a distal 5’ and 3’ duplex between nucleotides 12–15 of each strand, while maintaining the single-stranded ends bound in the observed active conformation. |
The internal cavity distribution allows for complete modelling of the RNA traffic inside the polymerase. This distribution implies that the template RNA enters the polymerase to be read and generates a double helix with the newly produced RNA. After a few nucleotides both RNA strands are separated, the nascent RNA exits the polymerase near the cap-snatching machinery locus and the template RNA exits the polymerase proximal to the entry channel. The template RNA can then be reincorporated into the RNP with minimal disruption to the macro assembly structure (Figures 1 and 3). This illustrates how the bunyavirus polymerase has adapted to work in the context of circular RNPs, and maintain their viral RNA as a single strand after duplication. The same mechanism is found in the polymerase structures of influenza virus and related nsNSV such as vesicular stomatitis virus, therefore the conclusions revealed by the LACV polymerase structure can be extended to all NSV polymerases that work in an RNP context (e.g. VSV, Ebola, Rabies, etc.) (Figure 1).
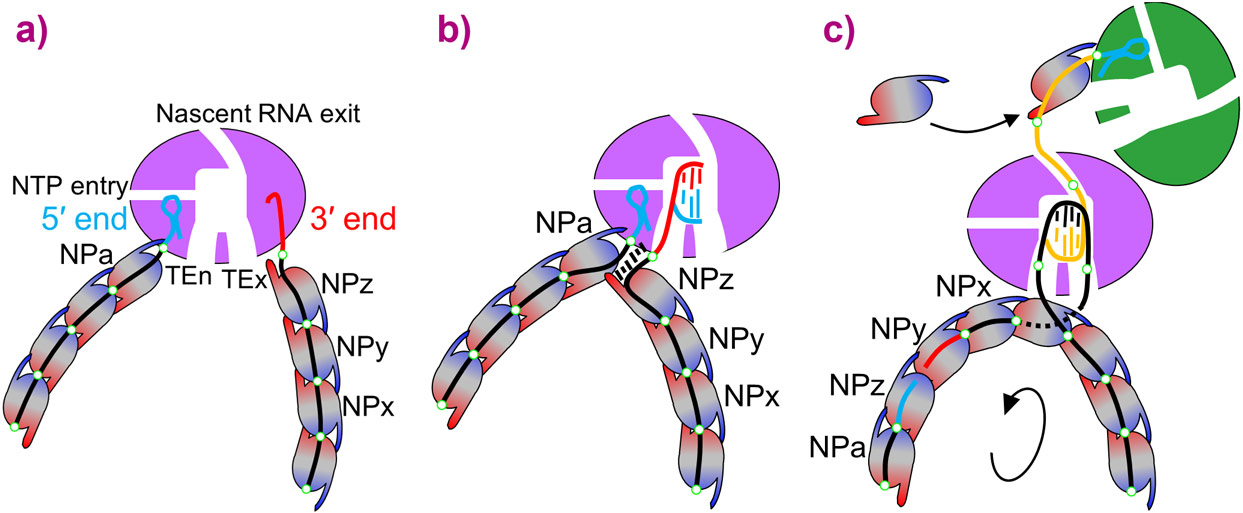 |
|
Figure 3. Proposed replication model. a) In the inactive state, whether after vRNP assembly or in virions, both ends of the genomic RNA are sequestered into the specific 5’ and 3’ RNA binding sites, thus circularising the RNP. b) With the 5’ end bound to the allosteric site for activation of RNA synthesis, a nascent cRNA begins to be synthesised. c) Once the nascent c50 end emerges from the exit channel it can recruit an incoming apo-polymerase as the first step in encapsulating the progeny cRNP with incoming apo-NPs. This may be facilitated by polymerase dimer formation [5]. |
In summary, based on these and previous studies [6], we propose a structure-based model of replication that might be extended to all sNSV and other related viruses such as VSV, Rabies or Ebola, which have a similar polymerase internal cavity and replicate in the RNP context (Figure 3).
Principal publication and authors
Structural insights into bunyavirus replication and its regulation by the vRNA promoter, P. Gerlach (a,b), H. Malet (a,b), S. Cusack (a,b) J. Reguera (a,b), Cell 161, 1267–1279 (2015) .
(a) European Molecular Biology Laboratory, Grenoble Outstation (France)
(b) Unit of Virus Host-Cell Interactions (UMI 3265), University Grenoble Alpes-EMBL-CNRS (France)
References
[1] R.M. Elliott, Nat Rev Microbiol 12, 673-685 (2014).
[2] J. Reguera, S. Cusack, D. Kolakofsky, Curr Opin Virol 5, 7-15 (2014).
[3] J. Reguera, F. Weber, S. Cusack, PLoS Pathog 6, e1001101 (2010).
[4] A. Pflug, D. Guilligay, S. Reich, S. Cusack, Nature 516, 355-360 (2014).
[5] B. Liang, Z. Li, S. Jenni, A.A. Rahmeh, B.M. Morin, et al. Cell 162, 314-327 (2015).
[6] J. Reguera, H. Malet, F. Weber, S. Cusack, Proc Natl Acad Sci U S A 110, 7246-7251 (2013).
Top image: Model of the LACV polymerase elongation state showing the trajectories of template RNA (cyan), product RNA (orange) and NTPs through the polymerase tunnels (green).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.