- Home
- News
- Spotlight on Science
- X-ray structure...
X-ray structure of malarial sugar transporter PfHT1 reveals the molecular basis for sugar specificity
22-06-2020
X-ray crystallographic studies reveal how the malarial parasite sugar transporter is able to transport many different sugars so effectively. Increased understanding of this molecular mechanism reveals the functioning of small molecule transports in general and could lead to new ways to selectively inhibit sugar transport.
Glucose transporters (GLUTs) work to maintain blood glucose levels in humans within a narrow range of concentrations (~ 4 to 12 mM), and do so with very high turnover rates (kcat of up to 6,500 molecules/s) [1]. At these physiological substrate concentrations, the GLUTs have relatively low affinities for the sugars, and yet somehow still maintain high specificity, i.e., not even stereoisomers of the sugars are transported. The molecular basis for how GLUTs can achieve such stringent selectivity with poor binding affinities is still poorly understood.
Typically, GLUTs have evolved to transport either D-glucose or D-fructose sugars with robust kinetics, but not both. Although crystal structures of the dedicated glucose transporter GLUT3 [2] and dedicated fructose transporter GLUT5 [3] were previously determined, it was still unclear, based on their sugar-binding sites, how their respective selectivity was achieved. Interestingly, a distant homologue from the malarial parasite Plasmodium falciparum hexose transporter 1 (PfHT1) was reported to transport both sugars efficiently, a likely advantage to the parasite [4]. Thus, the research described here aimed to crystallise and solve the structure of PfHT1 to understand how this feat is accomplished. X-ray diffraction data from PfHT1 crystals was collected at the beamlines ID30A-3 and ID23-1.
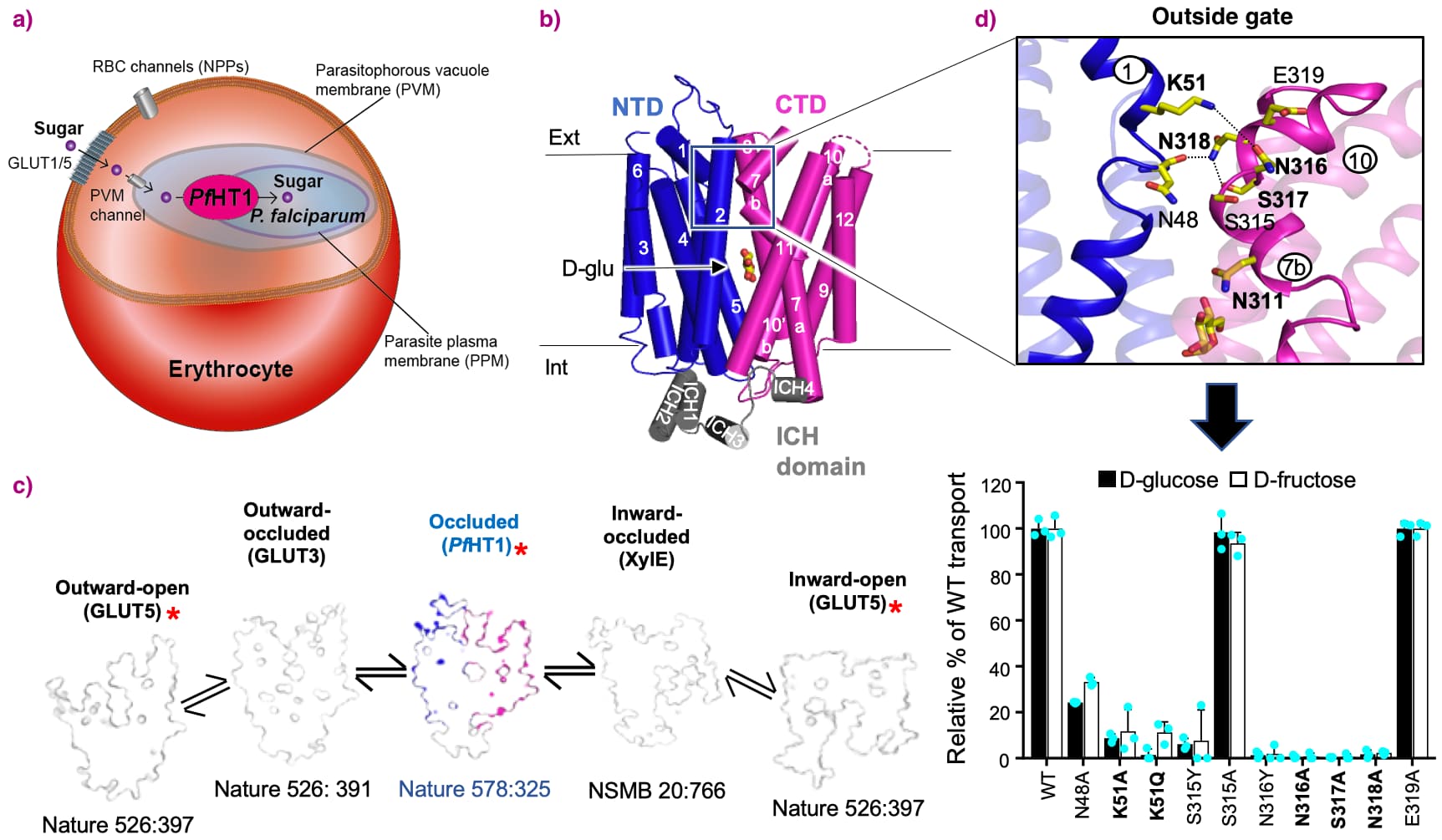
The first surprise was that the PfHT1 sugar binding site is almost identical to human GLUT3 [2]. Secondly, a symmetrical occluded D-glucose bound conformation had been captured (Figure 1), which was only thought to be transiently occupied during the GLUT transport cycle (as currently shown in textbooks). This unique state was significant and enabled two important mechanistic conclusions to be reached after inclusion of transport activity assays and MD simulations: (i) glucose transport in PfHT1 and related GLUT proteins must be driven primarily by conformational selection and (ii): PfHT1 has not evolved the sugar binding site to enable the transport of many different sugars, but rather it has evolved its gating dynamics to gain this competence.
 |
|
Figure 1. a) P. falciparum infected erythrocyte illustrating the localisation of PfHT1 within the parasitic plasma membrane and the GLUT-dependent uptake of hexoses, mainly D-glucose and D-fructose. b) The structure of PfHT1 in complex with D-glucose in the occluded conformation. PfHT1 is shown as cartoon, with the N-terminal six-transmembrane (6TM) domain (NTD) (blue), the C-terminal 6TM domain (CTD) (magenta), the intrahelical domain (ICH) (grey) and D-glucose shown as sticks. c) The fully-occluded conformation of PfHT1 (centre) was the last remaining state to be observed within the rocker-switch alternating-access mechanism of MFS transporters belonging to the sugar-porter subfamily. The observed structural states are shown as surface transversal cross-sections and from left to right: outward-open rat GLUT5 (PDB 4YBQ), outward-occluded human GLUT3 (PDB 4ZW9), fully occluded PfHT1, inward-occluded XylE (PDB 4JA3) and inward-open bovine GLUT5 (PDB 4YB9); red asterisk refers to structures determined from the Drew group. d) PfHT1 outside gating interactions between TM7b (magenta) and TM1 (blue), shown as cartoon (top) and indicated by dashed line. Critical and non-critical residues in the gating on TM7b and TM1 are highlighted in red/black respectively. Relative transport activity for TM1–TM7b interacting-residue mutants, bottom, for [14C]D-glucose (black bars) and [14C]D-fructose (white bars). |
Put simply, rather than evolving the sugar binding site to transport many different sugars, PfHT1 has evolved its outside gate to shut more easily and so become less stringent in which sugars bind at the sugar binding site. Indeed, mutations made up to 15 Å from the binding site were found to either abolish transport or had a reduced kinetic preference for D-glucose, and had shifted it towards D-fructose. From a broader perspective, this work highlights that gating dynamics might be more important for fine-tuning substrate preferences than previously thought. Indeed, it has been shown by work from Dorothee Kern and others that enzymes can shift their substrate preference by evolving dynamics [5]. Such mechanistic insight has pharmacological implications in the structural-based design of allosteric modulators or inhibitors of sugar transporters, which are drug target for diseases such as cancer, diabetes, or in the case of PfHT1, malaria.
Principal publication and authors
Qureshi A.A. (a), Suades A. (a), Matsuoka R. (a), Brock J. (a), McComas S.E. (a,b), Nji E. (a), Orellana L. (a), Claesson M. (a), Delemotte L. (b), Drew D. (a), The molecular basis for sugar import in malaria parasites, Nature 578, 321-325 (2020); doi: 10.1038/s41586-020-1963-z.
References
[1] Mueckler M. & Thorens B., The SLC2 (GLUT) family of membrane transporters, Molecular aspects of medicine 34, 121-138 (2013).
[2] Deng D. et al., Molecular basis of ligand recognition and transport by glucose transporters, Nature 526, 391-396 (2015).
[3] Nomura N. et al., Structure and mechanism of the mammalian fructose transporter GLUT5, Nature 526, 397-401 (2015).
[4] Joet T., Eckstein-Ludwig U., Morin C. & Krishna S., Validation of the hexose transporter of Plasmodium falciparum as a novel drug target, Proc Natl Acad Sci U S A 100, 7476-7479 (2003).
[5] Henzler-Wildman K. & Kern D., Dynamic personalities of proteins, Nature 450, 964-972 (2007).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.