- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2011
- X-ray imaging
- Visualising and interpreting the brain of Australopithecus sediba
Visualising and interpreting the brain of Australopithecus sediba
Evolution of the modern human brain from that of our last common ancestor with chimpanzees has seen a radical volume increase in the human lineage. Humans have evolved an average brain volume about four times that of chimpanzees, and have among the largest brains relative to body size of any living animal. In addition to these size differences, crucial organisational differences distinguish human and chimpanzee brains. Relative priority and timing of these critical processes in the evolution of the human brain – size increase and cortical reorganisation – have been debated since the discovery of the genus Australopithecus early in the 20th century [1].
The consensus opinion is that an australopith species gave rise to our own genus, Homo, between 2.3 and 1.8 million years ago. Au. sediba has been dated to slightly less than 2.0 million years, thus fitting firmly within this period. The reconstructed endocranial cast of Au. sediba (endocasts are proxies of brains estimated from the inner surface of crania) is surprisingly small (420 cm3), being only approximately 40 cm3 larger than average chimpanzee endocasts. Even compared to chronologically older australopithecines from South Africa (e.g., Au. africanus), the Au. sediba endocast is smaller. Such a small brain volume from a relatively late example is inconsistent with the notion of gradual brain enlargement amongst australopiths and across the transition to Homo. The small volume also is surprising given derived Homo-like morphologies exhibited by Au. sediba postcranial bones (e.g., hand and pelvis).
Comparing organisational details of endocasts requires high resolution surface details. In order to acquire images of appropriate resolution, we employed phase contrast X-ray synchrotron microtomography. Since the fragile fossil was embedded in matrix and far exceeded the maximum imaging size for a rock sample at the ESRF, we used a complex acquisition protocol on beamline ID17 that was developed for high quality imaging of large fossils. The protocol permits acquisition of high quality scans despite very low transmission of X-rays through the sample (less than 0.02%) even when the theoretical optimum is approximately 11%. Based on these exceptional data, we produced an endocast rendering from the Au. sediba cranium with visualisation of surface details exceeding most other hominin endocasts (Figure 137). Incorporation of computer-based techniques for in silico fossil preparation aided production of the rendering without necessitating destructive preparation [2].
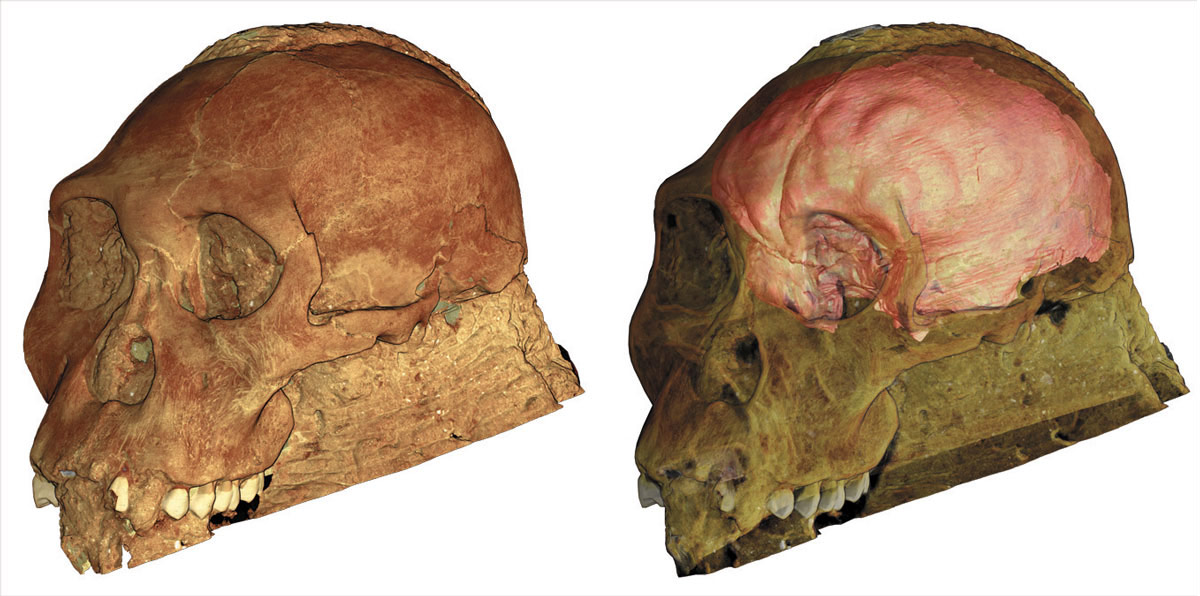 |
|
Fig. 137: Rendering of Australopithecus sediba cranium produced from images obtained on ID17 (left), with the cranium and surrounding matrix transparent to permit visualisation of the endocast inside (right). |
Comparing convolution patterns on the surface of the Au. sediba endocast to those of humans, other australopiths, and chimpanzees highlights similarities with the latter two groups (Figure 138). When examining features beyond overall size and convolution patterns, however, we observed proportional changes in the frontal lobe of Au. sediba that may foreshadow proportional changes modern humans exhibit relative to chimpanzees (Figure 138). For example, prefrontal cortex, particularly anteriorly, is relatively expanded in humans compared to other apes and is thought to support uniquely human capacities for multi-tasking, reasoning, planning, and innovation. The few South African australopithecine endocasts available for comparison with the Au. sediba endocast, all of which are attributed to taxa chronologically older than Au. sediba (e.g., Au. africanus), exhibit similar proportional changes, but to a lesser extent than the Au. sediba endocast. Based on these observations, and if Au. sediba is a human ancestor, we find support for gradual neural reorganisation of the orbitofrontal region in the transition from Australopithecus to Homo, but little support for gradual brain enlargement before the transition to Homo.
 |
|
Fig. 138: Top panel illustrates similarly-scaled human, chimpanzee, Australopithecus sediba and Au. africanus endocasts in inferior view. Note orbitofrontal organisation in the Au. sediba endocast resembles the human condition more closely than do the Au. africanus endocasts. Bottom panel illustrates same endocasts in left lateral view. Note similarities in surface configuration of australopithecines and chimpanzees. |
Principal publication and authors
K.J. Carlson (a,b), D. Stout (c), T. Jashashvili (a,d,e), D.J. de Ruiter (f,a), P. Tafforeau (g), K. Carlson (f) and L.R. Berger (a,h), Science 333, 1402-1407 (2011).
a) Institute for Human Evolution, University of the Witwatersrand, Johannesburg (South Africa)
b) Department of Anthropology, Indiana University, Bloomington (USA)
c) Department of Anthropology, Emory University, Atlanta (USA)
d) Georgian National Museum, Tbilisi (Georgia)
e) Anthropological Institute and Museum, University of Zürich (Switzerland)
f) Department of Anthropology, Texas A&M University (USA)
g) ESRF
h) School of Geosciences, University of the Witwatersrand, Johannesburg (South Africa)
References
[1] D. Falk, Yearb. Phys. Anthropol. 52, 49-65 (2009).
[2] A. Val, K.J. Carlson, C. Steininger, J.M. Kibii, C. Churms, B.F. Kuhn and L.R. Berger, S Afr. J. Sci. 107:11/12, (2011).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.